細菌可以演化成人嗎?進化論以親緣關係樹解釋萬物起源,但演化親緣關係樹的最基部的主幹竟是斷裂的?我們人類與地球上的生物最起初是從哪類生物演化而來的呢?
演化論學者藉『單一起源跨類演化』的親緣關係樹呈現各類別生物的演化關係,嘗試以DNA、RNA或蛋白質序列分析的相似性尋找分子生物學的演化證據,探尋原初地球上原始生命的共同祖先……
但由於各演化學者探尋的角度不同,彼此的證據浮現了更多演化理論的矛盾!依照演化原理,地球上生命的基本類別到底要分為三個類群(兩小一大:真細菌域、古菌域與真核生物域【依此觀點我們算是真核生物域生物之一】),還是兩個類群(一小一大:真細菌域、古菌域【依此觀點我們算是古菌域生物之一,德州大學學者甚至稱呼人類為『古菌人』】呢?目前演化論學者缺乏共識相持不下,公說公有理,婆說婆有理,吵翻天了!
目前生物學界在生物物種的分類是以演化親緣關係由低等至高等的樹狀圖概念予以分類的。目前主流學術界所採用的生物分類系統,則是由演化論者作為「系統分類學家」(Systematists)的學術共識所主導與決定的。現代分類學的核心原則是演化親緣關係學(Phylogenetics),主要透過化石證據、分析DNA、RNA、蛋白質序列相似性或形態特徵,利用統計與計算模型建立各類生物跨種演化的「親緣關係樹」,來視覺化呈現物種的共同祖先及分支脈絡(註1)。其根本邏輯就是:「萬物都是演化而來的;因此分類必須反映各類生物演化歷史(單系群)」。這代表一個生物分類階層只要無法呈現廣進化上的演化親緣關係,在現代主流分類學中就會被視為「不合格」(例如:將鳥類與哺乳類皆歸納為『內溫動物(體溫較為恆定)分類群』的分類方式因主流的演化概念(羽毛恐龍化石)牴觸,就被判定為不符合演化原理的『多系群』。)至於合格還是不合格,則是要看這個分類群合不合乎演化學者的『主流共識』所裁定的演化途徑!
依演化觀點的「五界系統」(Five-Kingdom System)由美國學者懷塔克(Robert Whittaker)於 1969 年提出。該系統依據細胞構造(原核/真核)、細胞數目(單細胞/多細胞)以及營養方式(光合作用/分解吸收/攝食)將所有生物分為五大界,為目前生物學基礎教育中最常見的分類方式:
1.原核生物界(Monera):細胞中心為環狀DNA的原始擬核(Nucleoid),
無核膜,多為單細胞,代表生物為:細菌、藍綠菌(藍綠藻)。
2.原生生物界(Protista):細胞中心為真正的細胞核(Nucleus;有核膜(內含不同數目的線狀DNA),多為單細胞或簡單多細胞,代表生物為:草履蟲、變形蟲、眼蟲(以上為單細胞),以及大型藻類(多細胞,如海帶、紫菜)。
3.菌物界(Fungi):細胞中心為真正的細胞核,多為多細胞 (酵母菌除外),細胞壁含有幾丁質。無法行光合作用,靠分泌酵素分解至細胞外界產生有機物後再吸收 (異營吸收)。代表生物:蕈類(如香菇)、酵母菌、黴菌。
4.植物界 (Plantae):細胞中心為真正的細胞核,全數為多細胞的真核生物,細胞壁含纖維素。能利用葉綠體行光合作用自製養分 (自營)。代表生物:苔蘚植物、蕨類植物、裸子植物 (如松、杉、柏)、被子植物 (亦即開花植物)。
5.動物界 (Animalia):細胞中心為真正的細胞核,全數為多細胞的真核生物,細胞無細胞壁。具備運動能力,主要靠攝食來獲取養分(異營攝食)。代表生物:各類無脊椎動物(如刺絲胞動物的水母、節肢動物的昆蟲、蝦蟹)、脊椎動物(如脊椎動物的魚類、鳥類、哺乳類)等。而我們人類則歸屬於『動物界』的『脊索動物門』其中的脊椎動物(即以脊椎骨保護脊髓)。
直到1977年卡爾‧沃斯(Carl Woese)提出以『原核生物拆分』的觀點去把分類生物分類,後來也演變成「六界」的分類方式。依據核糖體RNA(rRNA)序列的差異,沃斯氏將懷塔克的原核生物界再增為兩個界而總共成為六界:可分為真細菌界、古細菌界、原生生物界、真菌界、植物界和動物界。沃斯氏在1990年再進一步將所有的生物再增加一個更高的階層:『域』,因此生物分類的八個階層由上而下分別是:域、界、門、綱、目、科、屬與種,域則共分為『三大域』,於1990年後獲得演化學者普遍接受(註2;圖2-4):
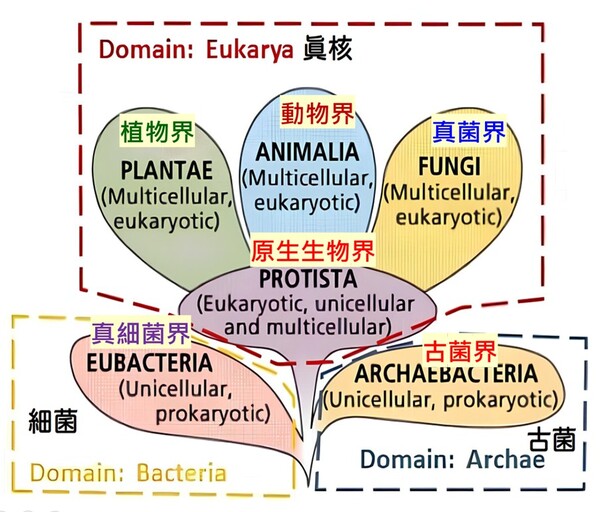
圖2-4 1990年後演化學者較普遍接受的三域六界系統分類圖,摘自咀嚼抹茶部落格 https://blog.gjlmotea.com/2082/%E7%94%9F%E7%89%A9%E7%89%A9%E7%A8%AE%E7%9A%84%E5%88%86%E9%A1%9E/
2.古菌域 (Archaea):僅含一個古菌界,由真正的古菌組成,單細胞原核生物,但在基因結構上被演化論學者認為與真核生物更具親緣關係,常生存於極端環境,包含嗜熱溫泉菌、甲烷菌與嗜鹽菌。
3.真核生物域 (Eukarya):包含了所有擁有真正細胞核、膜狀胞器(如粒線體、葉綠體、內質網等)以及細胞骨架的單細胞或多細胞生物四個界(原生生物界、真菌界、植物界和動物界)。從微小的單細胞酵母菌,到巨大的藍鯨與神木,乃至人類自身,都屬於真核生物域。
三域說與二域說的棘手難題——細胞膜分子結構大不同的化學矛盾
真核生物域生物的四個界與真細菌域它們的細胞膜結構均是以『雙層甘油磷脂質』(甘油=丙三醇,具有三個碳原子)形成其細胞的重要基本架構(圖2-5左),而此種甘油磷脂質是以第3號碳原子連結磷酸基形成的2號碳『G-3-P手性』(甘油-3-磷酸手性,圖2-5右)。而古菌的細胞膜之構成材料的甘油2號碳則是『G-1-P手性』(甘油-1-磷酸手性,圖2-5右)。
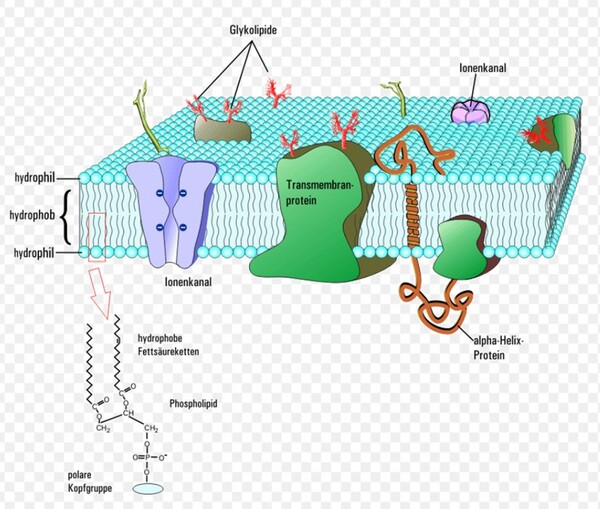
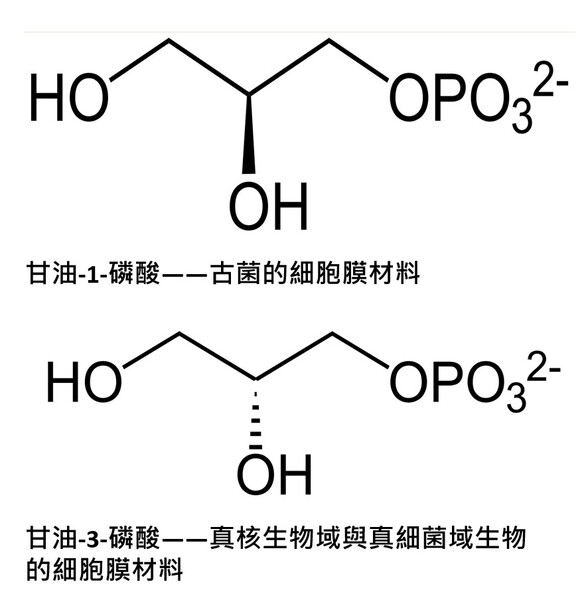
圖2-5(右) 在上半部(甘油-1-磷酸,Glycerol-1-phosphate,屬於古菌域細胞膜)的甘油磷酸分子結構圖中:2號碳(手性碳)的 OH 是黑色實心楔形(solid wedge)。在立體化學表示法中,黑色實心楔形代表鍵朝向觀察者,也就是「突出紙平面」。而在下半部(甘油-3-磷酸,Glycerol-3-phosphate,屬於真核生物域與真細菌域細胞膜)的甘油磷酸分子結構圖中:OH 是虛線楔形(hashed wedge)(凹入紙平面,遠離你)。另外兩條普通實線則表示位於紙平面內。圖片來源:維基百科關鍵字『細胞膜』,授權:CC BY-SA 3.0;『甘油-3-磷酸』授權:公有領域;英文關鍵字『Glycerol 1-phosphate』授權:公有領域。
1972年,美國加州大學的辛格和尼克森提出真核生物域細胞膜(真細菌域也相同)的流體鑲嵌模型(Fluid Mosaic Model)解釋細胞膜(或生物膜)的各種現象,是目前生物學界被最廣泛接受和認可的觀點。這種觀點主張:構成膜的蛋白質和雙層甘油磷脂類分子(甘油-3-磷酸結構)具有鑲嵌關係,而且膜的結構處於流動變化之中。隨後進行的研究有大量實驗結果支持流動鑲嵌模型的理論。在流體鑲嵌模型學說中,膜中的甘油磷脂質分子以雙層排列,構成了膜的基本結構。磷脂質分子為雙性分子,分為親水頭端和疏水尾端,雙層磷脂質分子之頭端(親水端)皆朝向細胞膜內側或外側的水相,因而使脂雙層分子之親水頭端面對細胞內與外,疏水尾端則兩兩相接埋於膜內。膜的另一種主要成分是蛋白質,蛋白質分子有的嵌插在脂雙層基本結構中,有的蛋白質貫穿細胞膜的兩側,有的則僅粘附於脂雙層的內外側表面上。這些蛋白質可能具有四種功能:1.運輸蛋白:跨膜的物質輸;2.酵素:協助細胞進行代謝;3.受體:與外界進行訊息溝通;4.辨識作用:辨別是否有「非自我細胞」入侵。
目前演化論學者對物種分類的看法上分成『三域說』與『二域說』兩大派系,這兩派其實都拿不出直接的化石證據,因此就致力發展DNA、RNA或蛋白質序列分析的相似性尋找分子生物學的演化證據,但是這樣的分子生物學分析在演化的不同學術機構的研究的不同解釋卻造成生物學界的『公說公有理,婆說婆有理』的『兩難』窘境。另有些時候不同演化途徑有時結果竟然分別演化出相似的生理結構(依演化原理不同演化分支應該生理結構也要不同)!這些窘境進化理論己乎都推給『趨同演化→因環境類似所以天擇演化出相似結構』這樣的說法:
1. 三域派 (Classic 3-Domain):由在1990年沃斯氏進一步提出,如以上所述:將演化親緣關係樹中所有生物物種都歸分於三大域的階層之下,即古菌域(Archaea)、細菌域(Bacteria)和真核域(Eukarya;即所有『有細胞核』的生物,包含動物界、植物界、真菌界與原生生物界)。進化論學者基於RNA資料的相似性,認為細菌、古細菌、真核生物三者的親緣關係如圖2-6所呈現的樹型關係:古細菌與真核生物擁有較晚近的共同祖先。
這一派遭受的困境是:無法解釋對於古菌域 (細胞膜構成為單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)為什麼其核心基因組與真核生物類似,但真核生物域細胞膜模式(雙層磷脂質、G-3-P手性;圖2-5)卻又演化倒退與更原始的真細菌域(也是雙層磷脂質、G-3-P手性;圖2-5)相同。最近的全基因組定序技術科學家發現真核生物的「資訊系統」(DNA 複製、轉錄)像古菌,而「操作系統」(代謝、脂質)卻像細菌。這使得三域假說遭遇無法確定演化分支的極大挑戰(註3)。因為根據華生與克里克的『分子生物學』基因表現的法則:DNA(基因組)→RNA(抄寫密碼)→蛋白質(決定遺傳表現),真核生物域與古菌域既有『相似的基因組』,為何細胞膜的遺傳表現會變得極端不同,且甚至是完全不同原理、不同性質的兩種細胞膜系統呢?
如果分成三個域在親緣關係樹的演化會產生演化論者最不喜歡的細胞膜為雙層磷脂質的多系群(Polyphyly):又稱複系群:亦即依某演化分類群包含的生物成員跨越兩個或多個主流演化分支,但這樣的分類群關係卻與三域說的主流演化分支(由三域說演化學者所討論的親緣關係學主流共識形成)牴觸。不同演化枝幹怎麼最後會演化出相同的結構呢?
多系群的歸類為『反主流』的演化概念!換句話說,就是『生物類群的共同特性』竟然與主流親緣關係樹牴觸!也就明明就是『不同分支(不同單系群)』的生物類群,依主流的演化原理應該是隨機突變與隨機環境選擇,演化出來的構造應該毫不相似,但彼此之間怎麼竟然還是彼此有極特殊的部分竟然是很類似的!這樣在進化論者上只好說是『趨同演化(convergence)』,亦即環境又變得很類似,所以生物又演化成很類似了。殊不知這樣的推論又可能把進化理論又帶入循環論證(都是你在講的)的風險之中。例如:硬骨魚與人類(後口動物演化分支)和烏賊(前口動物演化分支)依演化原理兩類生物理應截然不同,但若將牠們的眼球切開剖析,你會發現它們竟然都完美具備了相機型眼球(Camera-type eye)的構造:
- 剛好都有透明的角膜與虹膜(可以控制光線進入的量)。
- 剛好都有一個圓球狀的水晶體與懸韌帶(可以調節影像的焦距)。
- 剛好都有充滿感光細胞的視網膜(負責顯像)。
在嚴酷的海洋生存競爭中,要看清獵物和掠食者,這種「相機型」的幾何光學構造是最完美的物理學解法。於是,兩個完全毫不相干,分屬不同演化分支的物種,在相同的環境壓力下,竟然『無巧不成書』地各自摸索出了『同一套設計圖』!更何況廣進化親緣關係樹各類生物這樣例子還不少呢!然而若因此將烏賊的頭足綱與脊椎動物歸併為相機型眼球生物竟然要被視為多系群,違反主流演化共識。因此有學者指出:這樣看來,錯誤的可能是進化論本身那一套生物源自單一起源共同祖先,由簡趨繁的廣進化的假設!
約翰尼斯堡大學地質學博士凱特.拉希金(Casey Luskin)曾指出進化理論解釋的此種風險可能是肇因於進化論本身(單一起源共同祖先、由簡趨繁的概念)的假設(A廣進化成B)就是錯的(註4):
『當生物學家無法建構親緣關係樹時,他們常常會暫時訴諸其他過程來解釋那些不符合親緣關係樹狀模式的資料。其中一種解釋是『趨同演化』。演化生物學家假設生物在不同的演化譜系中獨立地獲得相同的性狀,而不是從共同祖先遺傳而來。每當演化生物學家被迫訴諸趨同演化時,這都反映出主要假設(指演化論本身)的失效,以及無法將資料擬合到樹狀模式。文獻中有很多這樣的例子,但幾個就足以說明(演化論本身有)問題了。』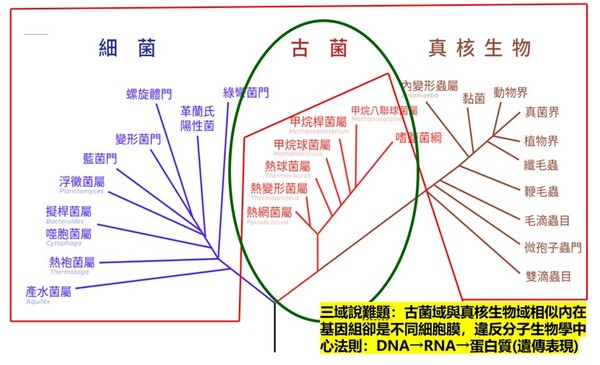
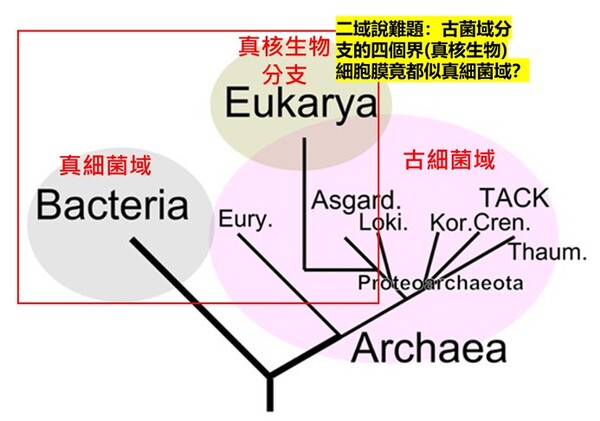
圖2-6 進化論三域派 (Classic 3-Domain;上圖) 二域派(2-Domain/Eocyte;下圖)與所遭遇細胞膜演化上的重大矛盾:紅色框內為細胞膜『雙層磷脂質,G-3-P,脂肪酸鏈』複系群分類生物;綠色框為細胞膜『單層/雙層甘油醚,G-1-P,具有異戊二烯支鏈』。
三域派的矛盾在於:為什麼真核生物的核心基因組【如同上游資訊系統】與古細菌類似,而細胞膜模式【如同下游操作系統】(雙層磷脂質、G-3-P手性、脂肪酸鏈)真核生物卻又趨同演化倒退到與真細菌(也是雙層磷脂質、G-3-P手性,脂肪酸鏈)相同。並且缺乏中間型態的化石。
二域派/融合派 (2-Domain/Eocyte): 認為真核生物是古細菌(阿斯嘉古細菌)的後代。但困境是無法解釋為什麼明明是不同的演化分支,細胞膜會在古菌(單層甘油醚、G-1-P手性、具有異戊二烯支鏈)進化為真核生物域的中途,細胞膜「砍掉重練」又被換回真細菌型(雙層磷脂質、G-3-P手性、脂肪酸鏈),並且缺乏中間型態的化石。
2. 二域派 (Classic 2-Domain): 二域系統是一種將演化親緣關係樹中的所有生物體分為兩個域的生物學分類:古菌域(包括真核生物)和真細菌域。它源自於對古菌多樣性認識的不斷深入,並對被廣泛接受的三域系統提出了挑戰。在1980年代,詹姆斯‧A‧萊克提出了始細胞假說,但由於當時的證據不足,該假說在很大程度上被三域系統所取代。21世紀初,人們對古菌,特別是它們透過與細菌共生在真核生物起源中所扮演的角色有了更深入的了解,這促使始細胞假說重新興起。特別是2017 年,日本科學家發現了一個名為普羅米修斯古菌(Promethearchaeati;舊稱阿斯嘉德古菌界Asgard archaea)的大型古菌界,似乎有進一步的證據表明,普羅米修斯古菌具有類似真核生物的蛋白結構(如RNA聚合酶與組織蛋白的相似性;膜變形與內吞作用的ESCRT-III 蛋白質系統),因此認為真核生物四個界應全數隸屬於『古菌域的分支』(註2)。科學家在 Asgard 古菌的基因組中,找到了大量過去被認為是真核生物特有的真核特徵蛋白質(ESPs, Eukaryotic Signature Proteins):細胞骨架(Cytoskeleton): 它們擁有編碼肌動蛋白(Actin)的基因,這能讓細胞改變形狀、長出突起。古菌的遺傳核心機制(DNA 複製、RNA 轉錄與蛋白質轉譯的分子機器),其與真核生物的相似度遠高於細菌。也使用一種類似組蛋白(Histones)的結構來纏繞包裝 DNA。依此觀點,在2023年甚至德州大學的論文戲稱我們人類乃是『阿思嘉德古菌人(Asgardians)』(註5)。
雖然基於細胞結構內部蛋白質的相似性,二域系統認為我們這些真核生物是古菌域的演化分支(即阿斯嘉德古菌是真核生物祖先的假說),但其演化上的困境是:無法解釋為什麼明明是不同的演化分支,為何細胞膜會在古菌(單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)進化為真核生物分支的中途,整個真核生物的細胞膜被「砍掉重練」換成細菌型細胞膜(雙層磷脂質、G-3-P手性、脂肪酸鏈),並且缺乏中間型態的化石?細胞膜可是生物體非常重要的生命結構,其上還有數以千計的膜蛋白作為細胞的門戶,細胞膜又負責古菌與真細菌的能量代謝(筆者註:古菌與真細菌沒有粒線體)。如果親緣關係樹上,真核生物域只是從古菌域演化出來的一個分支,明明有相似的遺傳核心機制,但為何其細胞膜的主結構化學成分(雙層磷脂質、G-3-P手性、脂肪酸鏈)卻反而與『另一個域』(真細菌域)極相似,而不是與古菌域相似呢 (如圖2-6) ?換言之,二域說的演化途徑仍是擺脫不了為何古菌域的『真核生物分支』其細胞膜最後卻演化為與另一真細菌域相同的『雙層磷脂質細胞膜』的嚴重矛盾(與二域說的主流演化分支牴觸)!
我們目前看到的 Asgard古菌(如 MK-D1),全身上下都還是百分之百的古菌醚鍵膜。它們體內雖然有真核生物的蛋白質(ESPs),但膜的生化結構卻沒有半點要「變成真核生物」的跡象。
這也是為什麼許多抱持批判思維的學者認為,單憑「基因相似度」就直接斷言人類是古菌人,有些過度簡化了細胞生化演化的複雜度。要跨越這道『脂質大峽谷』,需要的不是隨機突變,而是兩套完整且不相容的生化微工廠是否進行了完美的交接。但是目前這樣的交接在化石與現存生物都還沒看到一點蛛絲馬跡!
真細菌與古菌細胞膜與細胞壁結構矛盾『水火不容』顯示『生物並非單一起源』
儘管二域說在進化理論發展上逐漸佔上風,但進化論學者在科學論文中仍坦承二域說這其中細胞膜與細胞壁結構在演化過程如何由單一原始生物起源逐漸演變過程幾個大的嚴重難題:
1. 兩者生物膜物理特性極不相容,不可能混合存在:古菌域細胞膜 (單層/雙層甘油醚為基本構成)與和真細菌域細胞膜 (雙層磷脂質)兩種『磷酸甘油骨架』手性(chiraty)截然不同,很難混合成穩定的生物膜而變成混雜的四不像,因特性不明確,最後在功能上一事無成。進化論者卡弗里歐坦承(註6):
『細菌和真核生物的膜脂由脂肪酸透過酯鍵與甘油-3-磷酸(G3P)連接而成,而古菌的膜脂則具有異戊二烯基烷基支鏈,這些烷基鏈透過醚鍵與甘油-1-磷酸(G1P)連接,導致甘油磷酸骨架的立體化學結構相反。這種「脂質矛盾(Lipid Divide)」引發了人們對微生物生命演化的疑問,因為真核生物被認為起源於古菌,這需要膜組成發生根本性的改變。』
進化論者維克托.索喬(Victor Sojo) 在2014年的研究也指出(註7):
『古菌域和真細菌域是親緣關係樹最底層的分支。這兩個類群在形態上相似,並共享一些基本的生物化學特徵,包括遺傳密碼,但它們之間的差異卻十分顯著,堪稱生物學中尚未解決的重大難題之一。這兩個類群的細胞膜和細胞壁組成截然不同,而DNA複製機制似乎也毫不相干。……古菌域和真細菌域在碳和能量代謝、以及膜脂立體化學方面的巨大差異,反映了不同族群中主動運輸作用的兩種獨立起源。……它們在細胞膜和細胞壁、糖酵解、離子主動運輸甚至DNA複製等基本特徵上存在差異。
兩者細胞膜脂質的差異可能是解決生物學中這一重大未解難題的關鍵。古菌的磷脂側鏈通常是異戊二烯類化合物,而細菌的磷脂側鏈通常是脂肪酸。雖然這可能反映了適應性演化,但古菌和細菌在甘油磷酸頭基的立體化學結構上也存在差異。古菌脂質具有sn-甘油-1-磷酸(G1P)頭部官能基,而細菌則使用鏡像結構sn-甘油-3-磷酸(G3P)。目前尚無令人信服的選擇性解釋來說明這些相反的立體化學結構。而相關的酶,古菌中的甘油-1-磷酸脫氫酶(G1PDH)和細菌中的甘油-3-磷酸脫氫酶(G3PDH),在系統發育上也沒有相似性,表明它們是獨立起源的。』
2. 細胞膜如何在演化的過程結構大換血? 宿主古菌必須在進化為真核生物的某段時間之內,把自身的古菌細胞膜(單層/雙層甘油醚、G-1-P手性、具有支鏈異戊二烯鏈)完全替換成與原先完全不同的細菌細胞膜(雙層磷脂質、G-3-P手性、脂肪酸鏈),且在這個演替過程中細胞不能崩潰而死亡。但這聽起就像是神話故事。因為在更換細胞膜的演化過程中,稍有不慎,細胞就會因為滲透壓失衡或膜蛋白失效而直接解體。這還沒有考慮真細菌與古菌細胞膜上還有各種特殊的膜蛋白,兩類生物各自不同的酵素系統如何相容呢!進化論學者有試圖提出可能的演化過程的各種猜測,但是仍是屬於各說各話尚無定論的階段!兩種細胞膜結構矛盾,水火不容,誰演化成誰都不對,這樣豈不顯示『生物應非單一起源』呢?
3. 細胞壁為何彼此迴異? 古菌域與真細菌域兩種不只細胞膜非常不同,細胞壁相較兩者也是結構截然不同(圖2-9;註8)。舉例而言,古菌域的細胞壁完全不含真細菌域細胞壁的肽聚糖結構,而是S層(晶格狀蛋白質)或假肽聚糖外覆結構的細胞壁為主,而真細菌分革蘭氏陽性菌與陰性菌。陽性菌具有厚的肽聚醣外層細胞壁,有的陽性菌也會在外覆S層結構,革蘭氏陰性菌具有薄的肽聚醣外層與甘油磷脂雙層的外膜的複合細胞壁。這些截然不同的特徵都代表這兩種域的物種其實應該沒有演化關係。再者,革蘭氏陽性菌與革蘭氏陰性菌的細胞壁也是彼此大不同(圖2-9)!
-
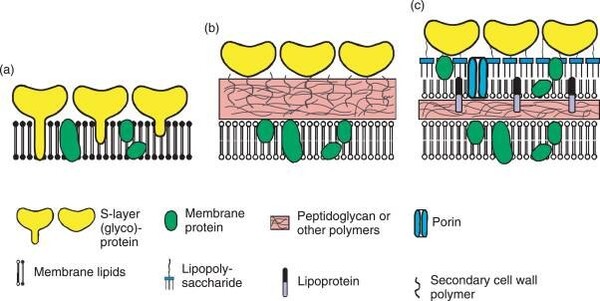 圖2-9 古菌與真細菌細胞膜與細胞壁比較圖
圖2-9 古菌與真細菌細胞膜與細胞壁比較圖 -
圖(a)某些古菌:具有甘油醚細胞膜與S層蛋白質外覆結構的細胞壁
圖(b)革蘭氏陽性菌細胞膜:具有厚的肽聚醣外層細胞壁,有的陽性菌也會在外覆S層結構
圖(c)革蘭氏陽性菌細胞膜:具有薄的肽聚醣外層與甘油磷脂雙層的外膜的複合細胞壁
圖片來源:https://boku.ac.at/en/btlw/nabi/research/generation-and-characterization-of-artificial-archaeal-cell-envelope-structures-and-their-relevance-as-model-membrane-platforms
表2-1古菌與真細菌細胞壁比較表
比較項目
古菌 (Archaea)
革蘭氏陽性菌 (G+)
革蘭氏陰性菌 (G-)
主要骨架成分
變異極大。多數為 S層蛋白質(S-layer),少數為偽肽聚糖(Pseudopeptidoglycan雙醣骨架為 beta-1,3 糖苷鍵)、複雜多醣或硫酸化多醣。
肥厚、多層的肽聚糖(Peptidoglycan雙醣骨架為beta-1,4 糖苷鍵)
薄、單層的肽聚糖(Peptidoglycan雙醣骨架為beta-1,4糖苷鍵)
肽聚糖厚度
無肽聚糖
厚(約20–80 nm)
薄(約 2–7 nm)
外膜 (Outer Membrane)
無
無
有(具有不對稱的脂雙層,外側含內毒素 LPS)
特殊化學成分
偽肽聚糖含 N-乙醯塔羅糖胺糖醛酸 (NAT) 且不含 D-胺基酸。
磷壁酸 (Teichoic acid) 與脂磷壁酸 (Lipoteichoic acid)
脂多醣 (LPS, 內毒素)、孔蛋白 (Porins)、周質空間 (Periplasmic space)
444.革蘭氏陽性菌與陰性菌外膜演化理論之爭:關於革蘭氏陽性菌(單膜、厚壁)與陰性菌(雙膜、薄壁與外膜)誰先演化出來?這兩類真細菌在化石與分子生物學的證據都非常缺乏演化的中間型。目前科學界仍有兩大主流假說,爭論的原因是革蘭氏陽性菌與陰性菌兩者「穩定的中間型難以存在」,兩派學者爭執不下:
假說一:陰性菌先演化出來(主張基因丟失的雙膜起源說) 一些學者(如 Thomas Cavalier-Smith)認為,最早的細菌具有外膜(類似陰性菌)(註9)。後來,某個分支因為遺失了製造外膜的基因,為了補強防禦,轉而加厚其肽聚糖層,演化成了陽性菌。這種「基因遺失」通常是快速且決定性的,不存在外膜「褪色到一半」的中間型。
假說二:陽性菌先演化出來(主張胞吞作用的單膜起源說) 另一派學者認為,最早的細菌只有一層膜與厚壁(註10)。外膜的出現,可能是早期某種胞吞作用、或是細胞壁外層脂質化後的突變產物(例如由芽孢的構造演變而來)。在這種模型中,外膜的獲得需要一套完整的脂多醣(LPS)與孔蛋白系統支撐,不完全的外膜結構在物理上是不穩定的,很難在自然選擇中存活下來。
5.原初生物膜結構作為模板的『結構遺傳』:加州大學細胞分子生物學博士約拿單.威爾斯(Johnason Wells)更震撼地指出表觀遺傳學的新觀點(註11):生物代代相傳必須直接繼承原本母細胞的「現成生物膜結構」作為模板。這種不依賴DNA序列、而是靠現有細胞結構傳承資訊的現象,在生物學中被稱為「結構遺傳(Structural Inheritance;註12)或「皮層遺傳」(Cortical Inheritance)。換句話說,古菌與真細菌的子細胞新細胞膜與新細胞壁無法僅僅依賴它們的DNA「從頭(de novo)合成」!
約拿單.威爾斯(JohnasonWells)指出:DNA 的確編碼了所有合成脂質的「酵素」,也編碼了細胞膜上的「蛋白質」。但是,DNA 沒辦法決定這些脂質和蛋白質在空間中要怎麼「排列」成一個雙層膜的球體。原因在於:
(1)脂質膜三維幾何資訊的缺失:DNA是一維的線性資訊(A、T、C、G 的排列)。當細胞內的酵素合成了新的磷脂質分子後,這些脂質分子並不知道細胞的「邊界」在哪裡、不知道該往哪裡擺放。這是物理熱力學的限制。換句話說:雖然脂質在水中會因為疏水效應自動聚集,但它們頂多只能形成雜亂無章的微膠粒(Micelles)或小脂質體,但脂質無法自動組裝成一個直徑數微米、具有特定極性、維持細胞內外滲透壓的巨大細胞膜。
因著母細胞生物膜的「模板(Template)」作用,新合成的脂質和膜蛋白質,必須「嵌入」現有的母細胞膜中。現有的膜就像是一個正在施工的建築框架,新的材料只能一片一片往這個框架上貼。
(2)膜蛋白的靶向(Targeting): 細胞核轉錄翻譯出膜蛋白後,這些蛋白質必須透過特殊的信號肽(Signal peptides),精準地尋找並插入現有的細胞膜(或內質網膜)。如果沒有那層現有的膜作為「靶標」,這些蛋白質就會在細胞質中亂成一團、失去功能。
(3)膜的擴展與分裂:當細胞生長時,新的脂質分子源源不絕地融合進母細胞膜,讓母細胞膜像吹氣球一樣脹大。最後,在細胞分裂時,母細胞膜從中一分為二。在這整個過程中,生物膜的「連續性(Continuity)」從未中斷過。 生物學界有一句名言: "Omnis membrana e membrana" —— 所有膜皆來自膜。這意味著從30億年前的第一個原始細胞開始,這條「膜的物理鏈條」就從未斷裂過,所有現存生物的細胞膜,都是遠古祖先細胞膜不斷擴展、分裂的產物!
6.細胞壁同樣需要模板的結構遺傳:細胞壁也是如此嗎?威爾斯氏指出:細菌和真菌的細胞壁同樣高度需要依賴現有結構。以細菌的肽聚醣(Peptidoglycan)細胞壁為例,它是一個巨大的、像網袋一樣包裹整個細胞的共價鍵分子(Sacculus)。當細胞要長大時,它不能把舊的牆拆掉重蓋,否則細胞會因為內部巨大的滲透壓而直接炸開。
細胞的作法是:先由特定的酵素在母細胞舊的肽聚醣網上精準地剪開一個小洞。緊接著由另一組酵素將新合成的醣鏈「編織」進這個缺口中。這種「一邊剪、一邊織」的精密工程,必須以舊的細胞壁網格作為物理藍圖母本。如果把細菌的細胞壁徹底去除(變成原生質體 Protoplast),即使它的DNA完全完好、合成細胞壁的酵素都在,這隻細菌也往往再也無法重新蓋出下一代子細胞正常的細胞壁!
約拿單.威爾斯(JohnasonWells)認為:達爾文主義過度簡化了生命,以為只要有 DNA 突變就能演化出新物種。但細胞膜的結構資訊是「非基因體資訊(Non-genomic information)」,這證明了生命在最源頭時,就必須預先具備一個結構完整、三維空間配置好的細胞原型。這種「DNA資訊與細胞原型同時存在」的作用自動反駁了化學進化論,也自動驗證了智慧設計論!
綜合以上所述,廣進化的親緣關係樹的基幹是嚴重斷裂且無法銜接的。古菌 (Archaea)、革蘭氏陽性菌 (G+)、革蘭氏陰性菌 (G-)三類生物分子生物學結構天差地遠,水火不容,演化論解釋無論推論誰演化成誰都不合理,同時化石證據與現存生物也都無法找到它們的中間型生物。這些現象其實強烈牴觸廣進化『生物單一起源』的樹型演化概念。這樣的結果強烈顯示:『生物應非單一起源』、『三個域彼此沒有演化關係』。以智慧設計論的觀點而言:若智慧設計者在其創造萬物時,預先制定『三個域』的生物細胞與生理結構的基本架構原本就彼此廻異,這樣若演化論者以達爾文『單一起源』的樹型演化概念予以強行解釋,結果自然格格不入了,您說是嗎?
- §參考資料
1.維基英文關鍵字:Phylogenetics,親緣關係學,又稱系統發生學
2.維基中文關鍵字:三域系統
3. Williams, T. A., et al. (2013). A reticulate view on the phylogenetic tree of life. Nature Reviews Genetics.
4.Casey Luskin, Problem 7: Convergent Evolution Challenges Darwinism and Destroys the Logic Behind Common Ancestry. Science & Culture Today網頁專文,February 9, 2015
https://scienceandculture.com/2015/02/problem_7_conve/
5.‘We’re All Asgardians’: New Clues about the Origin of Complex Life, June 14, 2023 • by Marc Airhart All complex life, a.k.a. eukaryotes, trace their roots to a common Asgard archaean ancestor.
https://cns.utexas.edu/news/research/were-all-asgardians-new-clues-about-origin-complex-life
6.Caforio, A., & Driessen, A. J. (2017). Archaeal phospholipids: Structural properties and biosynthesis. Nature Reviews Microbiology.
7. bioenergetic basis for membrane divergence in archaea and bacteria., PLOS Biology August 12, 2014, https://doi.org/10.1371/journal.pbio.1001926
https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.1001926#references
8. Generation and characterization of artificial archaeal cell envelope structures and their relevance as model membrane platforms, Institute of Synthetic Bioarchitectures, Boku university網頁專文
https://boku.ac.at/en/btlw/nabi/research/generation-and-characterization-of-artificial-archaeal-cell-envelope-structures-and-their-relevance-as-model-membrane-platforms
9. Cavalier-Smith T. Rooting the tree of life by transition analyses. Biol Direct. 2006 Jul 11;1:19. doi: 10.1186/1745-6150-1-19. PMID: 16834776; PMCID: PMC1586193.
10. Lake, J. Evidence for an early prokaryotic endosymbiosis. Nature 460, 967–971 (2009). https://doi.org/10.1038/nature08183
11. Jonathan Wells, Zombie Science: More Icons of Evolution, Chapter 6: DNA as the Secret of Life, p.111~134
12.英文維基百科關鍵字:Structural inheritance



