

目前進化理論基本上是認為『古代古菌(一種沒有細胞核的單細胞生物)』先吞噬真細菌形成其粒線體,之後演化為有核單細胞生物(如變形蟲),再由經過多細胞化的演化過程越來越複雜,形成動物、植物等多細胞的『真核生物』。但『自然界沒有古菌吞噬真細菌的任何先例啊!』,『生物的細胞核如何演化產生的呢?』這些問題多年來在進化理論中一直是難解的謎!以致在尋求演化的軌跡上一直也因著前寒武紀嚴重缺乏可佐證的化石出現,因而呈現演化軌跡與機制眾說紛紜、莫衷一是的局面。
甚麼是『古菌』呢?與一般的真細菌有甚麼不同呢?
過去曾經將古菌(Archaea)和真細菌一同歸為原核生物(沒有細胞核的單細胞生物),並將其命名為「古細菌(Archaebacteria)」,但這種分類方式已過時。事實上古菌被進化論者認為有其獨特的進化歷程,並與其它生命形式有顯著的生物化學上的差異,所以現在將其列為演化理論親緣關係分類系統—三域系統—其中的一個域。在這個系統中,古菌域、真細菌域與真核生物域各自為一個域,並進一步劃分為歸屬於其下的更低的七個階層—界(僅有古菌界)、門、綱、目、科、數與種。
古菌與真細菌有很多相似之處,即它們『環狀摺疊DNA』、『沒有細胞核而是擬核結構』與『沒有任何其他膜狀胞器』,同時另一些特徵則相似於真核生物,例如存在DNA重複序列與DNA環繞組織蛋白的核小體。古菌的細胞壁為醣蛋白、假肽聚醣或蛋白質;真細菌細胞壁則含肽聚醣。古菌和細菌的大小和形狀非常相似,但儘管如此,但進化理論認為古菌與真核生物的親緣關係更為密切,特別是在一些代謝途徑(如轉錄和轉譯)有關酶的相似性(如引子酶或DNA聚合酶)上。古菌還有一些性狀是獨一無二的,比如由依賴醚鍵構成的單層甘油醚細胞膜,而真細菌與真核生物則為雙層甘油磷脂質細胞膜。與真核生物相比,古菌的細胞膜有更多途徑的能量來源,從熟悉的有機物糖類(如使用特殊的Entner-Douderroff途徑分解醣類)、氨、金屬離子或氫氣(H2)還原二氧化碳以生成甲烷的代謝,多數古菌極端厭氧。真細菌則藉細胞膜使用糖解作用與有氧呼吸作用,有需氧菌、厭氧菌與兼性菌三類。古菌藉著二分裂法、出芽來進行無性生殖,但沒有發現能產生內孢子的古菌種類。真細菌則以二分裂法為主,某些菌種可形成內孢子對抗缺水、高溫或化學壓力環境。
最初古菌被認為都是一些生活在溫泉、鹽湖之類極端環境的生物,但近來也發現它們的棲息地其實也十分廣泛,從土壤、海洋、到河流濕地。甚至也被發現在人類的大腸、口腔、與皮膚。尤其是在海洋中古菌特別多。目前沒有已知的作為病原體或寄生蟲的古菌,他們往往是片利共生或互利共生。如甲烷菌生活在人和反芻動物的腸道中幫助消化,產甲烷菌還被用於沼氣生產和污水處理。溫泉中古菌中的酶甚至能承受高溫和有機溶劑,常被生物技術所利用。而真細菌生存範圍則極為廣泛;生態角色則是生產者(自營)、分解者(異營)或消費者(異營)三者都有。 (以上資料摘錄自維基百科—古菌、真細菌)
我們是『古菌人』?新興進化理論—古菌竟是動植物與人類的演化先祖?
進化論者近幾年試圖提出古菌演化為真核生物(即所有『有細胞核』的生物,數量眾多,包含動物界、植物界、真菌界與原生生物界)的「代謝共營模型」(syntrophic model)來解釋演化論上一直捉襟見肘的『細胞核如何演化產生』的疑問。新的理論認為是不同的細菌之間因彼此相互的代謝需求互利而產生的緊密互助關係(互補性的營養交換,如提供H2或還原性代謝物),進化論者認為代謝共營的兩種細菌最終導致了物理上的融合與細胞結構的複雜化,也就是太古的阿斯嘉德古菌在代謝上的相互倚靠的合併,最終導致其被阿斯嘉德古菌內共生吞噬了某種α-變形菌成為有『粒線體』的『原始真核細胞』。這就如同A車與B車一直在一起開(相依為命),有一天突然『A進入B之內』合併為一台『A+B拼裝車』,兩個方向盤也自動去掉一個。換句話說,新的演化理論認為在此種共營的狀況下真核生物可能是阿斯加德古菌『一次』偶然的『內共生』嵌入了α-變形菌則演化成其粒線體,之後再內摺化細胞膜形成有兩層膜的細胞核,再進化成其他更複雜的真核生物。之後又進一步第二次內共生添加了藍綠菌變成葉綠體,從而演化出藻類與綠色植物。
近代2015年一些科學家成功培養出的海底熱泉生態系分離出來的一些阿斯嘉德古菌(Asgard Archaea;如普羅米修古菌 Prometheoarchaeum),顯示它們具有長而複雜的細胞突起與近似肌動蛋白與微管蛋白的蛋白質,通常與共生的消耗氫氣的真細菌或產甲烷古菌生活在一起並進行代謝交換。這支持了前述的共養模型 (Syntrophic model):演化論者認為真核生物的誕生可能始於古代阿斯嘉德古菌用其突起包圍了細菌夥伴而將其『一次性吞噬(內共生)』,最終將其納入菌體內演化為其本身的粒線體。阿斯嘉古菌的命名者以北歐神話為其命名,意義是洛基城堡附近發現的——洛基城堡是位於挪威和斯瓦爾巴群島之間的大西洋中脊上的黑色煙囪。2015年的演化理論認為新發現的阿斯嘉德古菌就是太古的原核生物,只等一次關鍵的內共生事件,就開啟了通往複雜生命(如人類)的大門。因此阿斯嘉古菌被進化論者認定是與真核生物親緣關係最近的原核生物。(註1)
換言之,在阿斯嘉德古菌被發現之前,簡單的原核生物(細菌、古菌)與複雜的真核生物(具有細胞核與膜狀胞器)之間因複雜度差異極大而存在巨大的演化鴻溝。阿斯嘉德古菌被認為在演化上填補了原核生物與真核生物之間的「缺失環節」。另外阿斯嘉德古菌的基因組中含有大量被認為是「真核生物特徵蛋白質(Eukaryotic Signature Proteins, ESPs)」的基因。演化理論認為這證明真核生物所需的許多「複雜化的工具箱(Complex toolkit)」(如極簡單細胞骨架蛋白質、內膜系統的控制基因)在真核生物出現之前,就已經存在於原始的古菌之中了。這甚至可能影響了原先琳·瑪格利斯(Lynn Margulis)自1990年所主張的三域六界的分類觀點。瑪氏認為生命被分為三個『域』:真細菌域 (Bacteria)、古菌域(Archaea)和真核生物域 (Eukarya;包含四個界:原生生物界、真菌界、植物界與動物界【含人類】)。而新演化觀點認為:真核生物並非與古菌平行的獨立分支,而是直接起源於阿斯嘉德古菌內部的其中一個分支,因此只需分為兩個域:真細菌域與古菌域。新的進化理論認為我們真核生物在本質上其實是「演化得非常複雜的一群古菌」,我們甚至可被稱為阿思嘉德人(Asgardians)。無疑地,這樣新的內共生起源理論的確讓一些的進化論者非常雀躍。(註1;註2)
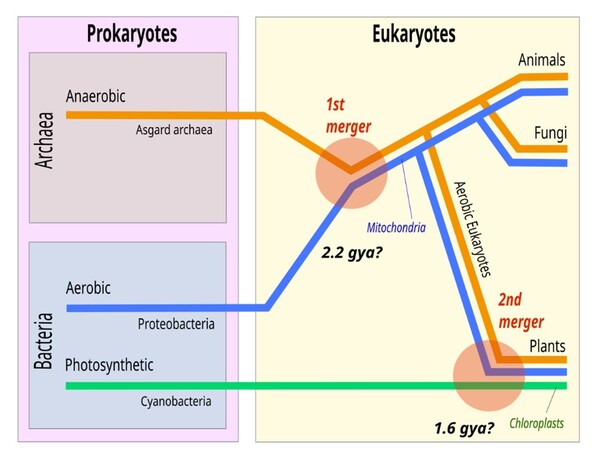
圖2-1 在新的共生起源論(Symbiogenesis)中,古細菌和需氧細菌的融合產生了真核生物,真核生物具有需氧線粒體;第二次融合又添加了葉綠體,從而產生了綠色植物。藍色線條代表加入粒線體的演化;綠色線條代表加入葉綠體的演化。圖片來源:Asgard Archaea 維基英文版CC BY-SA 4.0,Chiswick Chap從檔案:Symbiogenesis.svg重新繪製已反映新的進化理論。
阿斯嘉德古菌共養模型的嚴重問題其實顯而易見
儘管進化論學者投入了非常多的努力從阿思嘉德古菌與甲烷古菌的「共營模型」在尋找細胞核進化的蛛絲馬跡作為線索,但是深究進化論者探索廣進化合理化的邏輯,卻可能已經犯了一些致命的錯誤,或是刻意忽略、跳過一些顯而易見的巨大演化障礙。分述如下:
也就是說,要吞入一隻細菌之前,這隻古菌就需要先已經是進化完成具有細胞核與內膜運輸系統的真核細胞,才能使其具有吞噬作用的能力!但按演化論的推想演化為真核細胞具有吞噬能力理論上又是因為有粒線體(要先有粒線體產生好幾千倍的能量才能成為真核細胞,以應付其能量需求)才能演化出真核細胞。這不正是『A為真,因為B為真;而B為真,因為A為真』的循環論證(Circular Reasoning)嗎?這就告訴我們進化理論其實已經先敗給了它們自己自相矛盾不通的邏輯了!既然原先就不是真核細胞而只是一隻古菌,怎可以假設它『吞噬』了細菌形成粒線體?甚至最後還越長越大形成了真核細胞?(註6)正如務實派進化論者丹尼爾(Daniel B. Mills)所言:
『阿斯嘉德普洛米修士菌株MK-D1——形成膜支撐突出,具有獨特的分支圖案,與PhAT及早期類似模型相符。然而,MK-D1 顯然無法吞噬,因為它直徑僅 500 奈米,且缺乏演化與吞噬所需的蛋白質及能量產生方式。事實上,雖然阿斯嘉古菌及更廣泛的TACK古菌含有某些對吞噬至關重要且目前在其他古菌和細菌中尚未觀察到的蛋白質,但它們缺乏許多真正特有於真核生物的吞噬相關蛋白,以及最有可能來自細菌的吞噬相關蛋白質.因此,雖然肌動蛋白和微管蛋白對吞噬作用是必要的,但它們卻數量不足,這顯示古菌宿主細胞無法進行吞噬作用。』
『整體而言,儘管與真核生物最接近的古菌譜系中存在肌動蛋白、微管蛋白及其他對吞噬作用至關重要的蛋白質,但目前尚無證據顯示古菌中存在吞噬作用(或更廣泛的內吞作用)。』
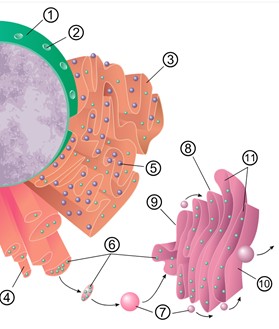
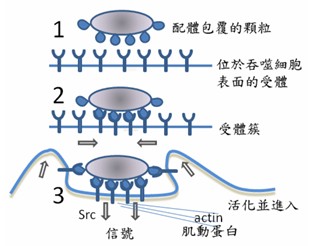
圖2-2真核細胞擁有內膜系統的運輸能力(左);吞噬作用如何以肌動蛋白(actin)進行(右)。內膜系統是真核細胞中由膜包被的細胞器(核膜、內質網、高基氏體、溶體、細胞膜等)組成的動態運輸網絡,透過「囊泡運輸」在各細胞器間進行蛋白質、脂質的合成、修飾與運輸。物質主要依循「細胞核外膜的內質網(生發囊泡) →運輸囊泡 →高基氏體→分泌囊泡→細胞膜」的路徑完成胞吐作用。 圖中標註:①細胞核 ②核孔 ③粗糙內質網 ④光面內質網 ⑤粗面內質網上的核糖體 ⑥囊泡中被轉運的蛋白質 ⑦囊泡 ⑧高爾基體 ⑨高爾基體順面 ⑩高爾基體反面 ⑪高爾基體囊泡產生處。而變形蟲藉由細胞膜上的受體「感知」周遭食物(如細菌、單細胞藻類)的化學訊號,當觸碰到食物時,細胞質流動伸出「偽足」(pseudopodia)包圍食物。這是一種非專一性的胞吞作用(phagocytosis),偽足在食物周圍融合向細胞內形成吞噬泡,隨後以肌動蛋白細胞骨架拉引此一吞噬泡進入其細胞質。而阿思嘉德古菌外有S-層形式的細胞壁阻隔,也不可能有變形蟲有細胞膜表面的受體去感知且包覆要吞噬的顆粒;也不可能事先有真核生物內膜系統的合作,怎麼會去做『吞噬作用』這件事來內共生呢?
再者,一般說來細菌(真細菌與古細菌)的細胞壁其實就會阻攔細胞本身去進行吞噬作用,因為細胞壁的存在就是為著形成外部堅硬結構方能保護細胞的。阿斯嘉德古菌和其他古菌一樣,其細胞壁缺乏肽聚醣,而肽聚醣卻是細菌普遍存在的細胞壁成分。阿斯嘉德古菌(大多數古菌亦同)膜外有一層由蛋白質亞基組成的剛性結構,稱為S-層(S-layer),提供物理保護。從這一點來看內共生理論的邏輯與古菌本身的生物化學結構是極端矛盾的!正如奧勒岡州州立大學的普通微生物學網頁還提到古菌與真核生物細胞壁特性的不同(註6):
『與細菌類似,古菌的細胞壁也是一種半剛性結構,旨在保護細胞免受環境和內部壓力的影響。』
如前所述,既然古菌的細胞壁有半剛性結構,進化論者難道還期望這樣的阿思嘉德古菌進行動物細胞或原生動物那樣的吞噬作用,內吞α-變形菌或甲烷菌嗎?目前科學家的共識是:阿斯嘉德古菌可能還不會「吃(吞噬作用)」,但是要命與矛盾的是:在演化論推論中它必須要先會『吃(吞噬作用)』,才可能演化出粒線體以便演化為真核生物。既然有這樣巨大的矛盾存在,阿斯嘉德古菌怎能算是廣進化「缺失環節」的重大發現呢?
2.阿斯嘉德古菌細胞膜結構(單層甘油醚)與細菌細胞膜(雙層甘油酯)兩者的主要差異極大(如圖2-3),若細菌被內吞演化為其粒線體將有極大的合併障礙。反而細菌的細胞膜較接近真核生物的細胞膜(雙層磷脂質)。阿斯嘉德古菌(單層甘油醚)之後若要演化為真核生物域(雙層磷脂質),兩者的細胞膜成分卻相差很多!古菌之後要接上真核生物域的演化矛盾必然會很大。有認真的演化論學者指出兩者細胞膜生物化學成分的六大不同點!說出這兩者生物膜的差異簡直大得嚇人(註6;註7):
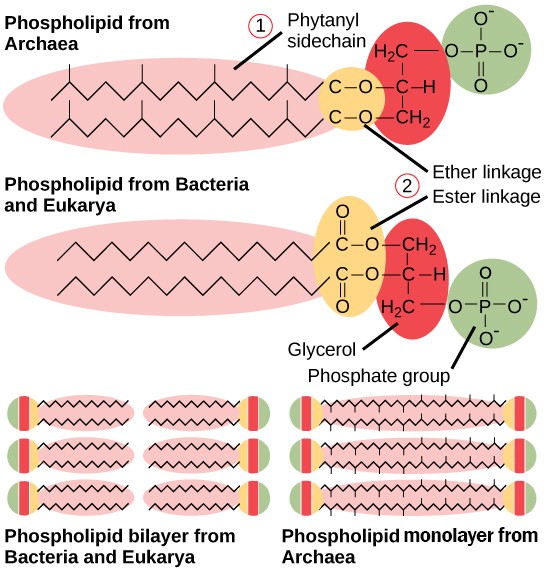
圖2-3摘自Opentax for AP課程生物學的圖22.15 古菌磷脂與細菌和真核生物的磷脂有幾個不同之處。首先,它們的植烷基側鏈是支鏈的,而不是線性的。其次,脂質與甘油之間是透過醚鍵而不是酯鍵連接的。第三,古菌的磷脂是單層結構;而細菌和真核生物的磷脂則是雙層的。
(1)脂質連接部位:阿斯嘉德古菌利用醚鍵(Ether-linkage)將疏水尾部與甘油連接,而細菌與真核生物則利用酯鍵(Ester-linkage)。
(2)尾部組成:阿斯嘉德古菌的脂質由支鏈異戊二烯(植烷基)組成,而細菌與真核生物則使用直鏈脂肪酸。
(3)甘油立體手性(Chirality):阿斯嘉德古菌細胞膜利用甘油-1-磷酸(G-1-P),而細菌與真核生物細胞膜則以甘油-3-磷酸(G-3-P)為基礎建構。2005 年由 Crauch 等人發表的論文(註9),其實觸及了一個非常根本的化學問題,這也是為什麼古菌與真核生物之間「沒有化石證據」時,科學家會感到如此焦慮。脂質矛盾的核心正是手性。這篇論文指出,細菌與古細菌的細胞膜不只是「化學鍵」不同(酯鍵與醚鍵),最麻煩的是甘油骨架的手性。細菌與真核生物: 使用的是 G-3-P (甘油-3-磷酸)。古細菌:使用的是 G-1-P (甘油-1-磷酸)。這兩者在空間上是互為鏡像的對稱分子。在生物化學中,合成這兩種不同手性分子的酵素(去氫酶)在結構上完全沒有關聯。這意味著:如果要從一種變成另一種,不是「突變」一下就好,而是要整套代謝系統砍掉重練(註7),難以用緩慢演化變異加以解釋。這正是二域論演化論學者所需面對的難題。
(4)膜結構:阿斯嘉古菌可以形成跨膜脂質單層(四醚結構),這使其在惡劣、通常高溫/酸性的環境中具有更高的穩定性,但這卻與大多數細菌與真核生物的雙層磷脂質結構不同。
(5)膜重塑蛋白:阿斯嘉德古菌擁有獨特的、複雜的膜重塑蛋白,如 ESCRT-III,它負責小囊泡形成和細胞分裂,雖然此一特徵更接近真核生物而不是細菌。但是阿斯嘉德古菌在吞噬細菌成為粒線體此點上勢必會形成二者不能相容的演化障礙。
(6)磷脂頭部和側鏈之間甘油鍵的手性:奧勒岡州州立大學的普通微生物學網頁還提到古菌與真核生物細胞膜上磷脂頭部和側鏈之間甘油鍵手性的不同(註8):
『古菌的細胞質膜具有一些獨特的特徵,使其與其他生物域區分開來。其中一個特徵是磷脂頭部和側鏈之間甘油鍵的手性。古菌的甘油鍵為左旋L-異構體( L-isomeric form),而細菌和真核生物則為D-異構體( D-isomeric form)。第二個差異是古菌的甘油鍵與側鏈之間存在醚鍵,而細菌和真核生物的甘油鍵則為酯鍵(Ester-linkage)。醚鍵賦予了細胞膜更高的化學穩定性。第三個和第四個差異與側鏈本身有關:細菌和真核生物的側鏈為非支鏈脂肪酸,而古菌的側鏈為異戊二烯鏈(isoprenoid chains)。這些異戊二烯鏈可以有支鏈側鏈。』
目前進化論認為真細菌較為古老。因此進化論者認為演化之順序是:真細菌→古細菌→真核生物。因為古細菌與真核生物在以下五方面特徵均較為相似。
(1)RNA聚合酶:古細菌的RNA聚合酶結構複雜,與真核生物的聚合酶類似;細菌則只有一種且構造簡單。
(2)轉錄因子:古細菌啟動基因轉錄所需的蛋白質,如TATA結合蛋白(TATA-binding protein),在真核生物中也能找到對應的版本。
(3)轉譯起始:兩者合成蛋白質時,起始的胺基酸通常都是甲硫胺酸(Methionine),而細菌使用的是甲醯甲硫胺酸(fMet)。
(4) DNA的內含子 (Introns): 雖然不如真核生物那麼普遍,但古細菌的基因中確實存在內含子,這在細菌中極為罕見。
(5)組織蛋白 (Histones): 這是最重要的特徵之一。古細菌擁有類似組織蛋白的蛋白質,用來繞並保護 DNA,形成類似「核小體」的結構。
但是奇怪的是:若將細胞膜成分之演化順序來比較,卻變成矛盾產生:真細菌(磷脂雙層)→古菌(跨膜脂質單層之四醚結構)→真核生物(演化倒退回到磷脂雙層)。進化論學者在2005年的科學論文中承認這是極大的矛盾,稱之為細胞膜的『脂質矛盾』(Lipid Divide;註9)。由單層脂質(古菌)倒退演化為原來『與細菌相同』的『兩層磷脂質』(真核生物)這樣的差異可不是小事!古菌單層細胞膜甘油醚(耐高溫卻不具柔韌性)結構也與真核細胞雙層甘油磷酯極為不同。雖然演化論學者還是有一套猜測的演化說詞加以解釋,但是他們卻不否認此一嚴重的矛盾確實存在,並且矛盾已經大到有此一專有名詞加以定名了。
3.細胞突起無助融合,無法解釋細胞體積為何增大數千倍:
|
特徵 |
阿斯嘉德古菌 (如 Lokiarchaeum) |
真核細胞 (如人體細胞、變形蟲) |
差距倍數 |
|
平均直徑 |
0.5−1.0μm |
10−100μm |
10 - 100 倍 |
|
估算體積 |
≈0.5μm3 |
500−50,000μm3 |
1,000 - 100,000 倍 |
進化論者認為阿
阿思嘉德古菌雖然體積小,但它擁有「長觸手」結構。如前所述,2020年日本科學家在《Nature》發表了世界首篇成功培養阿斯嘉德普羅米修士古菌(Prometheoarchaeum syntrophicum)的論文,發現了絲狀突起(Protrusions)的事實:這種古菌雖小,卻會長出非常長且細的分支狀突起。日本研究者認為這些觸手狀結構增加了表面積,讓它能與其他細菌(如未來的線粒體祖先)進行物質交換。進化論者認為體積的「暴漲」並非單純的球體變大,而是透過這些觸手將共生細菌「包裹」進來,最終合併成一個大型的複雜細胞。宿主細胞可能先演化出長長的絲狀結構來增加表面積,用以包裹住細菌(未來的線粒體)。一旦完成內共生,這些突起物收縮並融合,將細菌包在體內。這個過程會讓細胞體積在視覺上看起來迅速變大,但其實是結構的重排(註11)。
雖然日本科學家成功地分離出了這些阿斯加德古菌,似乎又為原核至真核的進化鴻溝提出了新的說法。然而極大不為人所知的問題隱藏在巨大掌聲的背後。作者承認:這些突起(Protrusions)非常細長且缺乏自主運動能力。它們不像真核細胞的偽足可以主動「獵食」,而更像是被動的「分支」。其物理瓶頸是這種突起缺乏真核生物那樣高效的肌球蛋白(Myosin)馬達(註11)。這意味著它們無法快速改變形狀,對於「抓取」移動中的物質效率極低,僅能與靜止或緩慢移動的共生細菌進行長期的「貼合」。
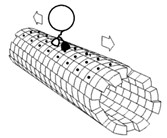
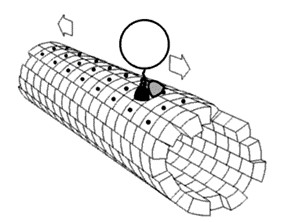
圖2-4 真核生物是以高效能的馬達蛋白(極高速走路拖著囊泡)消耗高能分子ATP輸送囊泡,所以在真核細胞雖空間大但輸送物迅速因此可以送達。在細胞內,最常見的馬達蛋白(如 Kinesin 驅動蛋白 或 Dynein 動子蛋白)在微管軌道上的移動速度通常為:平均速度約0.5至2 μm/sec(微米/秒)。某些特定的馬達蛋白在神經細胞中可以達到 5μm/sec(微米/秒)。如果把馬達蛋白類比成一個正常成年人的身高(約1.7 米),其運送速度相當於:每秒跑出100 到200米。時速高達360 到720 公里。這相當於在高鐵甚至飛機的速度下,還能精準地將「貨物」(囊泡)送到指定的房間。阿思嘉德古菌沒這樣肌球蛋白的輸送系統,細胞若變大1000~10000倍,卻僅能靠緩慢的擴散作用或細胞質巡流輸送物質,該準時送到的卻慢吞吞,細胞可能走向死亡。
同一年在另一篇論文的作者也殘酷地指出古菌細胞膜在演化過程中的結構弱點(註12):其化學限制是古菌的細胞膜通常由醚鍵脂類(Ether lipids)組成。這種醚鍵結構雖然耐高溫耐酸,但其柔韌性(Flexibility)與真核生物的雙層酯鍵脂類(Ester lipids)不同。因此這種膜結構在長出極細的突起時,極易受到環境應力(剪力)的破壞。這限制了細胞突起的長度與複雜度,若沒有穩固的細胞骨架支持,細胞突起會發生斷裂或塌陷。這些細胞膜化學分特性的限制因素其實都嚴重地限制阿思嘉德古菌(小細胞=原核細胞)不可能變大成為真核生物(大細胞=真核細胞)。
務實派的進化論者尼克連恩(Nick Lane)也多次強調指出:在沒有內共生體(粒線體)之前,產生任何突起都是「(能量)昂貴的負擔」(註13)。因細胞無端地突起將增加其表面積,也同時增加了細胞質的總體積。由於阿斯嘉德古菌是厭氧菌,無法進行主動運輸(Active transport)的能力,氧氣不易取得、ATP量少或代謝物在細長突起中的擴散速度極慢都成了演化過程中的致命傷。關鍵點是:如果突起過長,其末端會變成「代謝死角」,這將限制了阿斯嘉德古菌,使其無法無限制地擴大捕食範圍而形成真核生物。因此演化論者雖然發現了阿思嘉德古菌,但卻又產生了新的無法解釋的演化障礙。儘管阿思嘉德古菌擁有細胞突起這個新發現讓進化論者雀躍萬分,但這些新產生更嚴重的問題恐怕才是他們頭痛的開始!試問既有缺乏運動能力這些諸多強烈的限制前提,小籃球(阿思嘉德古菌)要如何攝入新的客體(變形菌—粒線體)內共生演化為一間大教室(真核細胞)呢?
4.無法跨越演化過程的巨大『能量鴻溝』:古細菌與真核生物在上述的五方面特徵均較為相似,但生物物理學家Nick Lane曾指出:問題在阿斯嘉德古菌的能量需求根本不足以演化成為真核生物(註13)!阿斯嘉德古菌是厭氧菌,使用極低能量就能存活,其對高能分子ATP的能量需求極低!依演化理論要適者才能生存!但為何卻往『能量需求極高且不利適應環境』的方向去演化呢?這是Nick Lane與其他的演化生物學家辯論最激烈的地方。也就是說,從一個「省錢度日」的節能厭氧古細菌,變成一個「揮金如土」的極度耗能的真核生物,這中間確實存在巨大的能量鴻溝。這個矛盾主要體現在以下層面:能量供應的「平方級」跳躍。細菌是靠細胞膜來產能的,若是體積增加1000倍,其細胞膜所產生的能量將極度不敷使用。因真核細胞的平均體積是原核細胞(古細菌或細菌)的 1,000 到 10,000 倍。如果只是單純把古細菌體積變大1000倍,它的細胞膜表面積(真細菌與古菌皆無粒線體)增長速度遠低於細胞體積急速增大數千倍(消耗能量的地方)。這將會導致細胞迅速因「能源短缺」而死亡,更何況阿思嘉德古菌還是能量需求極低的厭氧菌!正如尼克.連恩在書中所言(註14):
『技術上來說,體積愈大,表面積對體積的比例就越小。因為面積以平方增加體積以立方增加。長度變為兩倍則表面積會變為4倍(2×2=4),但是體積會變成8倍(2×2×2=8)。這會造成結果就是細菌長越大,能源效率就越差,因為用來產生能源的膜面積比起細胞增加的體積來說會變小。』
換句話說,既然阿斯嘉德古細菌在厭氧環境中利用氫氣或簡單有機物過活,產生的ATP根本少得可憐!它們根本沒有多餘的「能量資本」去養活複雜的細胞骨架、內膜系統和巨大的基因組。而相較之下真核生物是一種高度需氧且高度耗能的生物,其細胞內在生命無時無刻不需要極大量的ATP,尤其是原生動物與動物界生物的細胞。
因此從進化論者期待阿斯嘉德古菌進行『吞噬作用』的角度來看,能量的矛盾就是極大的問題!吞噬作用是細胞無細胞壁的動物巨噬細胞(白血球的一種,為真核細胞)等特殊細胞方能吞入大顆粒物質的『能量依賴型』過程。能這樣的進行吞噬的能量是因為真核生物有粒線體生產大量高能分子ATP。而進化論者又寄望古菌進行吞噬作用之後不將吞噬物消化而在細胞內形成『最初的粒線體』,這在邏輯上根本就是矛盾的!內共生寄望古細菌吞噬作用發揮功效,但古細菌若能吞噬作用,全程勢必依賴ATP高能分子水解提供能量。但大量的ATP高能分子水解提供能量卻又不是厭氧型(低耗能無氧代謝流程)且無粒線體的古菌能夠做到的,必需先有粒線體。這便是在邏輯上陷入循環論證的錯謬之中。更不用說吞噬作用的細胞骨架(肌動蛋白絲)根本與古菌的細胞骨架形式完全不同!
循環論證是種很好笑的『自己永遠說自己是對的』錯謬邏輯。譬如:認定「小孩就是要打」的某人,看到「不乖」的小孩會說:就是因為更小的時候父母親沒有打。但是後來又看到「沒被打但很乖」的小孩,他便會說:打了他會更優秀。
尼克,連恩在探討生物演化的『生命的躍升』這本書上有一段耐人尋味的文字,指出演化論學者所遭遇到這種古細菌演化成有核的真核細胞『能量的巨大障礙』的真實困境(註15):
『粒線體帶給細胞最大的厚禮,同時也是粒線體讓細胞可以快速演化的關鍵,就在於它們帶來早已準備好可以製造大量能量的內膜,以及整套可以就地調節呼吸作用的基因。只有當細胞裝備了粒線體之後,它才可能自我升級成為大而活躍的吞噬細胞,而免於因為過多能量消耗而伴手絆腳。如果上面推論都正確,那麼缺少粒線體的吞噬細胞(指內共生之前的原始細胞)應該從來不曾存在,因為沒有粒線體就不可能有吞噬作用。』
換言之,在演化論者的預測中在還沒有粒線體入駐前,阿斯嘉德古菌依然是個「窮光蛋」。但阿斯嘉德古菌演化為真核細胞的維持成本極高: 細胞膜的擴張(長出觸手)需要大量的脂質合成與能量。而能量根本入不敷出,而這是演化過程最致命的限制!這就如一家小公司永遠限定每個月『總電費絕不能超過1,000元,超過即予斷電』(如阿斯加德古菌是節能厭氧菌),這家公司在這樣的緊箍咒下,難道能期望它未來發展為台積電或輝達這樣的大公司嗎(演化為真核生物)?
5. 細胞突起運輸的物理流體動力學的限制(擴散障礙):阿斯嘉德古菌這些突起雖然細長,但它們極其脆弱且受限於化學分子的運動規律。在缺乏能量無法進行耗能的細胞主動運輸的情況下: 阿斯嘉德古菌雖然有類似真核生物的細胞骨架蛋白(類Actin結構),但在進化出成熟的「馬達蛋白」(如圖2-4耗能的肌球蛋白Myosin細胞輸送系統)之前,它無法像現代變形蟲那樣快速伸出偽足。阿斯嘉德古菌細胞突起內部的物質傳遞主要依賴布朗運動(細胞質循流與緩慢的擴散作用)。如果觸手長度超過一定限度(幾微米以上),營養物質從細胞突起的末端傳回細胞本體的速度會慢到無法支撐整個代謝,這當然會限制了它「抓取範圍」的極限。如以下幾個期刊所述:
(1)2017年尼克.連恩的論文提到細胞突起的末端將發生代謝停滯(註13):如果突起長度從1μm增加到10μm,營養物質到達細胞中心的時間會增加 100 倍。在缺乏真核馬達蛋白(如Myosin)進行「細胞質流」(Cytoplasmic streaming)的情況下,這會導致突起末端發生代謝停滯。
(2)2020年日本科學家今町洋之(Imachi, H.)的論文(註11)提到這些細胞突起極其細長(直徑僅約50-100μm),因此這種高曲率(Curvature)的膜結構在流體動力學上非常不穩定。研究者也發現這些突起是被動且僵硬的。因為缺乏強大的能量支撐來維持細胞內部的微管運輸,這些突起無法像真核生物的偽足那樣快速移動或進行「主動吞噬」。它們只能靜態地增加與共生細菌(如產氫細菌)的接觸面積,以克服擴散障礙帶來的吸收效率問題。
(3)2025年艾克爾與羅賓遜(Akıl, C. & Robinson, R. C.)在論文中提到了阿思嘉德古菌將遭遇「細胞質擁擠」(Macromolecular crowding)效應(註16)。在狹窄的突起中,大分子蛋白質的擴散係數(Diffusion coefficient)會大幅下降。如果沒有像真核生物那樣的「囊泡運輸」(Vesicle transport),細胞根本無法跨越這道物理障礙來維持大型細胞的運作。
由以上所述,阿思嘉德古菌細胞突起內擴散速度慢,將導致限制了突起長
度,細胞無法無限制變大。而且在獲得粒線體(如果真可以演化的話)前,這
樣的細胞突起複雜結構反而是「負資產(拖油瓶)」。第三,因為細胞膜突起
是甘油醚化學成分,結構容易斷裂,且難以進行快速的膜融合。以上這些輸
送因素對於向真核細胞的演化方向全是不利的!
6. 阿斯嘉德古菌是脆弱的共養型細菌:阿斯嘉德古菌既是供養型古菌,當然非常倚靠另一種細菌的存在而生存,本身生存非常嬌貴且弱不禁風。
阿斯嘉德古菌到底是脆弱嬌貴到甚麼程度呢?日本科學家今町洋之的研究團隊是從日本南海海槽(Nankai Trough)海拔2533公尺深的深海沉積物中取得,花了近12年才將其培育成功!前5年在生物反應器中進行富集培養,後續數年進行分離與純化。且阿斯嘉德古菌因屬厭氧菌,其生長極慢,倍增時間約為14 至25 天(而大腸桿菌僅需約20分鐘)。不只如此,實驗證實MK-D1還必須與產甲烷菌(Methanogen)或硫酸鹽還原菌共生,透過轉移氫氣或甲酸來代謝。
奇怪的是:阿斯嘉德古菌生長條件如此嚴酷,演化論者竟然要寄望它成為強勢且進化真核生物(強勢生物)的共同祖先,這是邏輯矛盾的!照進化理論不是應該要強勢物種才符合『適者生存』的叢林法則的嗎?我個人的判斷真正的原因可能是因為『找不到合適的人選』,蜀中無大將,只好找『廖化』當先鋒。這樣沒有魚『至少有一隻蝦』。
以上六項問題在原核細胞(古代古菌)進化為真核細胞(古代變形蟲)的過程都是十分嚴重的障礙!演化過程的化石證據也是嚴重地缺乏。因此古菌進化為真核生物的進化過程甚至可能根本就不存在!期盼媒體與教科書編寫者務必正反面平衡報導,若是一面倒不平衡的偏袒進化理論給予報導,恐有失客觀與公允,違反科學實事求是的精神了!
§參考資料
1. 維基中文關鍵字:阿斯加德古菌
2.Peter Rüegg, 生命的起源:微生物如何奠定了複雜細胞的基礎,蘇黎世聯邦理工學院網頁專文,2025年3月21日
3.維基英文版關鍵字:Asgard Archaea
4.維基英文版關鍵字:phagocytosis
5.維基百科關鍵字:內膜系統;吞噬作用
6.Daniel B., The origin of phagocytosis in Earth history, 12 Jun 2020, Interface Focus (2020) 10 (4): 20200019
7.普通微生物學(General Microbiology)網頁古菌Archaea篇,奧勒岡州州立大學(Oregon State University)
https://open.oregonstate.education/generalmicrobiology/chapter/Archaea/
8. 圖2-2摘自Opentax for AP課程生物學的圖22.15
https://openstax.org/books/biology-ap-courses/pages/22-2-structure-of-prokaryotes
9. Jonathan H. Crouch, Michael J. Russell, Simon G.W. Mullidge & David A. Hartley, The lipid divide: segregation of the three domains of life through subcellular compartmentalization?. Progress in Lipid Research, 2005, Jan. Vol. 44, Issue 1, p. 1–16.
10. 維基英文版關鍵字:Motor protein
11.Hiroyuki Imachi, Yusuke Nobu, Masaru K. Nobu, Nozomi Nakahara, Yuki Morono, Isolation of an archaeon at the prokaryote–eukaryote interface, Nature(2020. Jan. 15), Volume 577, Issue 7791, Pages 519–525.
12. Dey, G., et al. (2020). Asgard archaea and the origin of the eukaryotic cell. Nature Reviews Microbiology, 18, 73-88.
13. Nick Lane, Origin of the Eukaryotic Cell, Molecular Frontiers Journal, Vol. 1, No. 2, pp. 108–120, 2017.
14.尼克.連恩,生命的躍升—40億年演化史上最重要的十大關鍵】,p.157,梅苃仁譯,貓頭鷹出版社。
15.同上,p.157~158。
16. Akil, C., & Robinson, R. C. (2018). Genomes of Asgard archaea encode functional actin. Nature. 以及最新的預印本 Radler, P., et al. (2025). Dynamic protrusions mediate unique crawling motility in Asgard.
